
Dilophosaurus facts for kids
Quick facts for kids DilophosaurusTemporal range: Lower Jurassic
|
|
|---|---|
 |
|
| Reconstruction of a Dilophosaurus skull | |
| Scientific classification | |
| Kingdom: | |
| Class: | |
| Superorder: | |
| Order: | |
| Suborder: | |
| Family: |
Dilophosauridae
|
| Genus: |
Dilophosaurus
|
Dilophosaurus (meaning 'double-crested lizard') was a medium-large dinosaur from 190 million years ago.
Dilophosaurus has been examined several times over the years and has been put into no less than nine different theropod groups. It may be related to the smaller Coelophysis. Specific details of its life-style are few. Some of its fossils were found in terrain which was a low-lying slow-moving river system at the time. The animal was obviously a carnivore.
Contents
Description

Dilophosaurus measured about 7 metres (23 ft) long and weighed about 400 kilograms (880 lb).
The teeth of Dilophosaurus are long, but have a fairly small base and expand basally. Dilophosaurus had 12 maxillary teeth and as many as 18 dentary teeth; the teeth were smaller in the tip of the upper jaw. The second and third front teeth feature serrations, which are absent in the fourth. Another skull feature was a notch behind the first row of teeth, giving Dilophosaurus an almost crocodile-like appearance, similar to the putatively piscivorous spinosaurid dinosaurs. This "notch" existed by virtue of a weak connection between the premaxillary and maxillary bones of the skull. The braincase is well known in Dilophosaurus, and is significant in that it bears a feature of the top side wall that is absent in ceratosaurians. Compared with ceratosaurians, the distal scapular expansion in D. wetherilli is uniquely rectangular. The upper leg bone (femur) is longer than the lower leg (tibia).
A study by Robert J. Gay (2005) comparing various specimens found no indication that sexual dimorphism was present in Dilophosaurus, so males and females would have been largely the same in terms of skeletal anatomy.
According to Rauhut (2000), Dilophosaurus can be distinguished based on the following features: a lacrimal bone with a thickened dorso-posterior rim; cervical vertebrae that have neural spines with a distinct central "cap" and an anterior and posterior "shoulder"; a scapular blade with a squared distal expansion; the presence of thin, paired nasolacrimal crests extending vertically from skull roof, each with a fingerlike posterior projection (according to Carrano, 2012)
Cranial crests

The most distinctive characteristic of Dilophosaurus is the pair of rounded crests on its skull, made up of extensions of the nasal and lacrimal bones. These are considered to be too delicate for anything but display purposes. Dodson (1997) noted that cranial crests first appeared in Dilophosaurus and were later retained, in one form or another, by other theropods.
The function of the crests on the skull of Dilophosaurus have been the subject of speculation among scientists ever since they were discovered. Traditionally, these bizarre cranial structures (and similar structures and post-cranial armor in other dinosaurs) were thought to be variously for attracting mates, intimidating/fighting rivals in the group, and intimidating potential predators of other species. However, Padian, Horner and Dhaliwal (2004) argued that phylogenetic, histological, and functional evidence indicates that these bizarre structures were most likely used for intra-species recognition.
Discovery and species

The first Dilophosaurus specimens were uncovered by Sam Welles in the summer of 1942 in the Kayenta Formation in Arizona. The site had been found by a Navajo, Jesse Williams, in 1940. Three individuals were present but one of these was too weathered to make an excavation worthwhile. Two specimens were brought back to Berkeley for cleaning and mounting by the team of Wann Langston, the holotype UCMP 37302 (specimen on which a name is based), and a second skeleton, the paratype UCMP 37303. They were given the name Megalosaurus wetherilli in 1954 by Welles. The specific name honoured John Wetherill who had explored the area of the find.
Returning to the same formation in 1964 to determine from which time period the bones dated, Welles found a new specimen, UCMP 77270, not far from the location of the previous discovery. Based on the unique double crests preserved for the first time in the new skeleton, Welles realized that the species was significantly different from Megalosaurus. In 1970, he re-classified it in a new genus, which he named Dilophosaurus, from the Greek words di (δι) meaning "two", lophos (λόφος) meaning "crest", and sauros (σαυρος) meaning "lizard"; thus, "two-crested lizard". The available evidence strongly suggests that both the holotype of D. wetherilli and the second skeleton are juveniles.
In 2001, Robert Gay reported the discovery of two, or possibly three, new specimens of Dilophosaurus wetherilli in the collections of the Museum of Northern Arizona. The specimens were found in 1978 in the Kayenta Formation, in the Rock Head Quadrangle. Gay found the specimens to compare favorably with the holotype (UCMP 37302). Gay noted that this material is significant in that it includes parts of the pelvis not preserved in either the holotype or in the "D. breedorum" specimen UCMP 77270.
Gay reported that some elements in the MNA collection were representative of an infant specimen (MNA P1.3181). This included a partial humerus, a partial fibula, and a tooth fragment. The infant specimen is significant for several reasons; first, it lends insight into the growth rates of early dinosaurs, and second, it provides researchers with an opportunity to compare the growth rate of Dilophosaurus wetherilli with that of a related species like Coelophysis bauri which could provide insights into early theropod growth rates. This specimen is also the first known infant of the genus.

Hu Shaojin (1993) assigned specimen KMV 8701 to a second species, Dilophosaurus sinensis. This species was recovered from the Yunnan Province of China in 1987, with the prosauropod Yunnanosaurus. Compared to D. wetherilli, this species was larger and more robust. Lamanna et al. (1998b) examined the material ascribed to D. sinensis and found it to be synonymous with Sinosaurus triassicus. This finding was confirmed in 2013.
A proposed third species, D. breedorum, was coined by Samuel Welles through Welles and Pickering (1999) for the specimen UCMP 77270 collected in 1964. Tykoski (2005) performed an ontogenetic analysis in his thesis and concluded that D. breedorum is an adult Dilophosaurus wetherilli. This was the first specimen of the genus to preserve a nearly complete cranial crest. Welles concluded that differences between the original specimens and UCMP 77270 justified the naming of separate species. He was unable to complete a manuscript describing this during his lifetime, and the name eventually came out in a private publication distributed by Pickering. This species has not been accepted as valid in other reviews of the genus, which see it as a nomen nudum and/or a junior subjective synonym.
Paleobiology

Dilophosaurus is considered to have been an obligate biped based on the presence of long hindlimbs oriented vertically under the pelvis, and short forelimbs that did not support quadrupedal locomotion. The hindlimbs suggest a fast and agile runner, as would be expected in a carnivorous theropod.
Welles (1984) proposed that Dilophosaurus traveled in small groups, based on the fact that several individuals were found together. Gay (2001b) noted that there was no direct evidence for this and noted that "flash floods would pick up scattered and isolated material from different individuals and deposit them together in the same area". Cranial display features make sense in social, gregarious animals, where other members of the species are available to observe and interpret messages of sexual status.
Feeding
The presence and distribution of non-interdigitating sutures in the skull of some reptilian groups, including Dilophosaurus, has been interpreted as indicating the presence of a system of levers, driven by jaw muscles, as an aid to predation. Welles (1984) rejected this hypothesis and interpreted the potential mobility in the skull of Dilophosaurus as a sign of weakness, and argued that the loose connection of the premaxilla precluded the capture and subduing of prey. This led to the early hypothesis that Dilophosaurus scavenged off carrion, because its teeth were too weak to bring down large prey. Recently, some researchers have argued that Dilophosaurus may have been piscivorous, due to the presence of Eubrontes-sized swim tracks at some localities, as well as anatomical similarities with spinosaurids.
Growth
Tkach (1996) conducted a study of the bone structure of the type species and concluded that Dilophosaurus wetherilli could have attained growth rates of nearly 35 kilograms (77 lb) in early life, which reflects a rapid growth rate.
Paleopathology

One Dilophosaurus wetherilli specimen bears a damaged vertebra that may have been injured during life and a potential abscess on the individual's upper arm bone. Another specimen has an unusually small left humerus compared to a very robust right arm, a possible example of fluctuating asymmetry. Fluctuating asymmetry results from developmental disturbances and is more common in populations under stress and can therefore be informative about the quality of conditions a dinosaur lived under. A specimen of Dilophosaurus from the University of California Museum of Paleontology labelled as UCMP 37302 displays eight different pathologies to its pectoral girdle and forelimb bones, including three bone tumors, broken and re-healed bones on both arms, deformed digits and an inability to utilize one forelimb. Though it is not certain, it is believed that most or all of the injuries on this specimen were acquired in a fight or an accident and that the theropod would have been in severe pain as it healed. The pathologies of this specimen were documented in the journal PLOS One.
In a 2001 study conducted by Bruce Rothschild and other paleontologists, 60 foot bones referred to Dilophosaurus were examined for signs of stress fracture, but none were found.
Paleoichnology

In the Kayenta Formation in Arizona, the same formation that yielded the original specimens for this genus, the trackways known as Kayentapus hopii and Dilophosauripus williamsi were attributed to Dilophosaurus by Welles (1971). These ichnotaxa feature a series of three-toed footprints that are consistent with the expected size and shape of the feet of Dilophosaurus. In 1991, trackway specialist Gerard Gierlinski re-examined tracks from the Holy Cross Mountains in Poland and renamed them Kayentapus soltykovensis, concluding that the "dilophosaur" form was the most appropriate candidate for making these ichnotaxa.
Fossilized footprints, discovered in 200-million-year-old sedimentary rock, which were assigned to Dilophosaurus were discovered in the Höganäs Formation in Vallåkra, Sweden, during the 1970s. The footprints appears to show that these dinosaurs lived in herds. Fossilized footprints assigned to Dilophosaurus have also been discovered in Sala, Sweden. Other tracks discovered in the Höganäs Formation have been assigned to the ichnogenus Grallator (Eubrontes) cf. giganteus, which were discovered in Rhaetian strata, and Grallator (Eubrontes) soltykovensis, which were discovered in Hettangian strata. A few of the tracks were taken to museums, but most of them disappeared in natural floodings. In 1994, Gierlinski and Ahlberg assigned these tracks from the Hoganas Formation of Sweden to Dilophosaurus as well.

Gierlinski (1996) observed unusual traces associated with a track specimen in the collection at the Pratt Museum in Amherst, Massachusetts. Specimen AC 1/7 is a "dinosaur sitting imprint", made when a dinosaur is resting its body on the ground, leaving an impression of its belly between a pair of footprints. Traces associated with AC 1/7 were interpreted by Gierlinski as the imprints of feathers, suggesting that Dilophosaurus was a feathered dinosaur. Further analysis proved, however, that the lines that seemed to be feathers were in reality just cracks in the mud where the animal sat. While this does not rule out the possibility of feathery covering on this species, there is no evidence for it and it currently remains as speculation.
Tracks of Eubrontes and Gigandipus of the Connecticut River Valley, which have been found in both Connecticut and in Massachusetts, have often been attributed to Dilophosaurus, however no fossil remains of Dilophosaurus have been directly attributed to either one of the footprint types. The size and shape suggested they were made by a theropod around 20 feet long similar to that of Dilophosaurus, suggesting they were either made by Dilophosaurus or a very close relative. Two similar footprints, Anchisauripus and Grallator, also found in the valley, have often been attributed to Dilophosaur's smaller relatives, Podokesaurus and Coelophysis, and occasionally attributed to Dilophosaurus itself.
Paleoecology
The remains of the type specimen of Dilophosaurus wetherilli UCMP 37302 and partial skeleton UCMP 37303 were recovered in the Silty Facies Member of the Kayenta Formation, in northeastern Arizona. The remains were discovered in 1942 and 1964 in blue shale and brown/gray siltstone that was deposited during the Sinemurian-Pliensbachian stages of the Early Jurassic, approximately 196-183 million years ago. Two other specimens assigned to this genus were discovered in the same formation; one in 1982 (UCMP 130053) in channel sandstone and the other, a scapula in terrestrial sandstone.
The Kayenta Formation is part of the Glen Canyon Group that includes formations not only in northern Arizona but also parts of southeastern Utah, western Colorado, and northwestern New Mexico. It is composed mostly of two facies, one dominated by silty deposition and the other dominated by sandstone. The siltstone facies is found in much of Arizona, while the sandstone facies is present in areas of northern Arizona, southern Utah, western Colorado, and northwestern New Mexico. The formation was primarily deposited by rivers, with the silty facies as the slower, more sluggish part of the river system. Kayenta Formation deposition was ended by the encroaching dune field that would become the Navajo Sandstone. A definitive radiometric dating of this formation has not yet been made, and the available stratigraphic correlation has been based on a combination of radiometric dates from vertebrate fossils, magnetostratigraphy and pollen evidence. It has been surmised that the Kayenta Formation was deposited during the Sinemurian and Pliensbachian stages of the Early Jurassic Period or approximately 196 to 183 million years ago.

The Kayenta Formation has yielded a small but growing assemblage of organisms. Most fossils are from the silty facies. Most organisms known so far are vertebrates. Non-vertebrates include microbial or "algal" limestone, petrified wood, plant impressions, freshwater bivalves and snails, ostracods, and invertebrate trace fossils.
Explorations in the 1970s and 1980s by James M. Clark, Farish Jenkins and David E. Fastovsky and collection and analysis by William R. Downs have produced several vertebrate specimens. The Kayenta Formation has produced several mass burial sites, and the remains of three coelophysoid taxa of different body size, which represents the most diverse ceratosaur fauna yet known.
Remains assigned to Dilophosaurus were recovered in the Upper Member of the Dharmaram Formation, in Andhra Pradesh, India. The remains were discovered in mudstone that was deposited during the Sinemurian stage of the Early Jurassic, approximately 196-189 million years ago. The same sediments also contained the fossils of a crocodilian, a sauropodomorph and the sauropod Lamplughsaura dharmaramensis.
In popular culture
Dilophosaurus is featured in both Michael Crichton's 1990 novel Jurassic Park and its 1993 movie adaptation. It is depicted as venomous, being able to spit venom, aiming for the eyes to blind and paralyze its prey (much like a spitting cobra); in the film, it also has a retractable neck frill around its neck (much like a frill-necked lizard). There is no evidence to either prove or disprove either the frill or the venom spitting, which was acknowledged by Crichton as artistic license. In the film, Steven Spielberg also reduced the size of Dilophosaurus to 3 feet (0.91 m) tall and 5 feet (1.5 m) long in order to avoid confusion with Velociraptor. Dilophosaurus was designated as the state dinosaur of Connecticut.
Images for kids
-
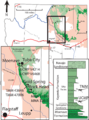
Map showing localities in northern Arizona where Dilophosaurus fossils have been collected (squares)
-

Life restoration showing hypothetical feathers and crest-shape
-
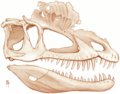
Reconstruction of the skull showing preserved extent of the crests
-

Cast of Coelophysis, with which Dilophosaurus has often been grouped, Redpath Museum
-

Reconstructed skeleton of the crested Cryolophosaurus, sometimes considered a close relative of Dilophosaurus
-
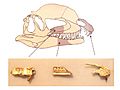
Skull bones of Dracovenator, which may have been a close relative of Dilophosaurus (onto whose skull the bones are plotted), ROM
-

Diagrams of Late Triassic-Early Jurassic tracks, several of which have been attributed to theropods similar to Dilophosaurus, including Eubrontes (A), Dilophosauripus (E), and Kayentapus (F)
-

Cast in Japan of a resting trace from Massachusetts, which was argued to have been made by a theropod like Dilophosaurus and to include feather impressions around the belly (arrow), but this has been questioned
-
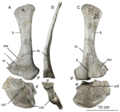
Scapula of Sarahsaurus with a probable tooth mark on its outer surface (C) possibly left by a scavenging Dilophosaurus
-

Restoration of Dilophosaurus in a bird-like resting pose, based on a resting trace at the St. George Dinosaur Discovery Site, Utah
-
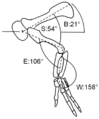
Diagram showing the forelimb in hypothesized resting posture
-

Reconstructed skull showing possibly outdated crest-shape, American Museum of Natural History
-

Restoration of an adult tending to its hatching clutch
-

Restoration of the right hand of the holotype in flexion, with the deformed third finger (below) unable to flex
-

Restoration of Dilophosaurus chasing Scutellosaurus
-

Models showing Jurassic Park style neck frills and outdated hand postures, Nong Nooch Dinosaur Valley
-

Restoration showing adult size














See also
 In Spanish: Dilophosaurus wetherilli para niños
In Spanish: Dilophosaurus wetherilli para niños
